







Происхождение, расселение и эволюция дуба.
Ископаемые остатки дубов широко распространены и были обнаружены на трех континентах. Анализ окаменелостей показывает, что дубы существовали еще 55 миллионов лет назад. Самое раннее появление дубов (Quercus) в летописи окаменелостей находится в палеоцене. Пыльцевые зерна дубов в позднем палеоцене (около 55 млн. Лет) в месте св. Панкраза (Австрия) в настоящее время являются самыми ранними свидетельствами рода (Hofman et al., 2011). А некоторые из самых ранних останков, найденных в Северной Америке (Bones, 1979; Manchester, 1994), Китае (Jiang, 1993) и Европе (Kvaček & Walther, 1989), относятся к эоцену. Богатое разнообразие дубовой пыльцы в древних отложениях указывает на то, что группы Quercus, Lobatae и Protobalanus, а также группа Ilex уже присутствовали в арктических районах среднего эоцена.
Почти одновременное появление рода на трех континентах, о чем свидетельствуют обнаруженные сегодня окаменелые останки, подняло вопрос о географическом происхождении и распространения рода. Два сценария были предложены, чтобы восстановить историю рода.
В первом сценарии (Zhou, 1992) род появился в Юго-Восточной Азии, происходящий от родственного рода Trigonobalanus во время палеоцена, и мигрировал в двух направлениях: в Европу и Америку через Североатлантический сухопутный мост (NALB) до эоцена, и через Берингов пролив после миоцена. Сценарий Чжоу (1992)
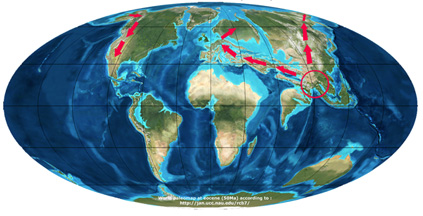
Во втором сценарии (Manos and Stanford, 2001 и Trelease, 1924) род Quercus произходит из широко распространенного бореально-тропического лиственного леса, который простирался по всему северному полушарию в начале третичного периода. Род далее дифференцировался, поскольку разделение между континентами стало более выраженным. В результате виды дуба появились на разных континентах в качестве разъединеной между Азией и Северной Америкой исконной группы, составляющей бореально-тропический лес. В Азии «наследственная часть» дифференцировалась на подрод Cyclobalanopsis и секцию Cerris рода Quercus. Виды группы Cerris мигрировали позже на запад в Европу. — В Америке «наследственная группа» дифференцировалась на секцию Lepidobalanus (белые дубы), секцию Protobalanus и секцию Erythrobalanus (красные дубы) с последующей миграцией белых дубов из Северной Америки в Азию и далее в Европу.
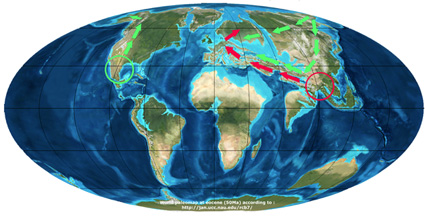 Scenario of Manos et al. (2001)
Scenario of Manos et al. (2001)
Хьюберт и соавт. (2014) предложили миграцию через Североатлантический сухопутный мост (NALB) и Берингию. И хотя данные окаменелостей не противоречат североатлантической связи, это говорит о том, что Берингов маршрут был, вероятно, закрыт на 12 млн лет (Barron et al., 2017). Ранее Manos и Stanford (2001) предположили, что время расхождения между белыми дубами Северной Америки и Евразии составляет 17 млн. Лет, что относительно согласуется с преобладающим представлением о раннем коридоре для растений и животных. Тем не менее, это не согласуется с общей низкими генетическими различиями между североамериканскими и евразийскими представителями белых дубов (Denk and Grimm, 2010).
В целом, объединенные данные современных филогенетических исследований и окаменелостей (палеоботанические и филогеографические результаты) показывают, что трансатлантический коридор для дубов существовал до позднего миоцена (Barron et al., 2017). И в любом случае, необходимо оценить потенциальное значение сухопутного моста для миграции рода Quercus между Европой и Северной Америкой во время раннего кайнозоя.
Несмотря на разногласия по поводу происхождения рода, палеоботаники соглашаются с чрезвычайно быстрым разделением рода во время олигоцена и миоцена в ответ на важные климатические изменения. Большинство ископаемых останков того периода похожи на ныне существующие виды. Следовательно, считается, что большинство сохранившихся видов существовало уже в середине миоцена (Trelease, 1924; Axelrod, 1983).
В целом, объединенные данные современных филогенетических исследований и окаменелостей (палеоботанические и филогеографические результаты) показывают, что NALB действовал как трансатлантический коридор для дубов до позднего миоцена (Barron et al., 2017). И в любом случае, необходимо оценить потенциальное значение сухопутного моста для миграции рода Quercus между Европой и Северной Америкой во время раннего кайнозоя.
Окаменелости Европы.
Род Quercus явно присутствует в Европе на переходе палеоцена – эоцена (около 55 млн. Лет) в Санкт-Панкраце (Австрия; Hofmann et al., 2011). Одним из немногих мест позднего палеоцена, который можно подтвердить, является Мен, Франция. Здесь листья были отнесены к Q. lonchitis Unger, Q. parceserrata, Q. proofctifolia Saporta или Quercus subfalcata (Barron et al., 2017). В эоцене окаменелости из рода Quercus широко распространены в Европе. Самые ранние свидетельства группы Ilex в Европе получены из пыльцевых зерен, найденных в открытом карьере в Коспудене (Германия) у таких эоценовых видов Quercus, как Q. rectinervis и Q. olafsenii, которые были распространены во флорах Евразии в середине позднего эоцена. играет важную роль в экосистемах. От олигоцена до среднего миоцена Quercus rhenana (возможно, раздел Lobatae) был доминирующим видом во многих местах и был разбросан по Центральной Европе, населяющей берега рек и болотистые районы (Barron et al., 2017) Виды группы Cerris (например, Q. kubinyii) являются важным элементом, населяющим европейскую флору раннего миоцена и неогена. Одним из наиболее актуальных дубов в Европе от среднего миоцена до раннего плиоцена является Q. pseudocastanea, широко распространенный в России, Армении, Казахстане и Западной Сибири. Точно так же Q. mediterranea является типичным цветочным компонентом во время неогена во всей Европе, широко распространенной до Европы, появляющейся в конце олигоцена. В предсредиземноморской растительности центральной и южной Европы ксерофитовые вечнозеленые дубы ( Cerris и Ilex) были важными элементами во время неогена. Некоторыми значительными видами для этого периода были Q. gigas, Q. pseudocastanea, Q. kubinyii, Q. mediterranea и Q. drymeja. Секция Cyclobalanopsis также присутствует в Европе (как C. stojanovii) во время плиоцена (Barron et al., 2017). В Северной Африке самые ранние древесные останки, относящиеся к роду Quercus, относятся к олигоценовому периоду, особенно Quercoxylon retzianum из Окаменевшего леса Каира (El-Saadawi et al., 2011). Одним из наиболее примечательных явлений раннего плиоцена является кольцевидный дуб, Cyclobalanopsis stojanovii в Болгарии (бассейн Бели Бряг). Из позднего миоцена наиболее распространенными видами в целом в Европе были Q. pseudorobur и Q. roburoides (Barron et al., 2017).


Азия
Одной из первых окаменелостей древесины Quercus является Quercus cretaceoxylon из верхнего мела в Японии (Suzuki and Ohba, 1991). Аналогично, первое появление Quercus как рода относится к раннему палеоцену Quercus tsagajanica в Поярке, Россия (Barron et al., 2017). Окаменелые листья, желуди и древесина Cyclobalanopsis из Японии и Китая были отнесены еще к эоцену, подобно Cyclobanalopsis naitoi в Японии (поздне-средний эоцен). древних окаменелостей в азиатских странах олигоцена мало, одним из наиболее важных является Quercus protoserrata, который похож на современные Quercus serrata и широко распространен в Восточной Азии (Barron et al., 2017). Недавно Xu et al. (2016) обнаружили много листьев окаменелостей циклобаланопсиса в поздних миоценовых слоях округа Мангканг, восточный Тибет, принадлежащие к Q. tibetensis (где до сих пор не сохранилось ни одного существующего циклобаланопсиса), что указывает на то, что дубы Циклобаланопсисы существовали во время позднего миоцена в центральной зоне Цинхай-Тибетского плато, и позволяет предположить, что климатические условия на юго-востоке Цинхай-Тибетского плато во время позднего миоцена, вероятно, были более теплые и более влажные, чем сегодня.
Расчетная палео-высота ископаемого участка Q. tibetensis во время позднего миоцена была, по меньшей мере, на 160 м ниже современной высоты, что указывает на продолжающийся подъем восточного Тибета со времени позднего миоцена. Поднятие юго-восточной части Цинхай-Тибетского плато подтверждается драматическими изменениями растительности в неогене: в позднем миоценовой флоре Kajun преобладает Quercus subgenus Cyclobalanopsis, в то время как поздняя плиоценовая флора богата видами секции Ilex, которая обычно встречается в субальпийском или альпийском районы юго-западного Китая. (Xu et al., 2014).
С раннего олигоцена группа Cerris распространилась и разделилась на множество видов в Евразии; однако, этаже секция вымерла в Северной Америке в неогене. С другой стороны, представители подрода Cyclobalanopsis (= Cerris, Denk et al., 2017) исчезли из Северной Америки, вероятно, в конце палеогена, а Европа — в конце неогена. Теперь этот подрод присутствует только в Восточной Азии.


Северная Америка.
Самыми древними найденными дубовыми желудями в Северной Америке является Quercus paleocarpa из среднего эоцена Кларно, росшие в Орегоне приблизительно 44 млн лет назад, который связан с подродом Cyclobalanopsis. Окаменелости из эоцена (особенно листья) скудны в Северной Америке, в частности из-за трудностей в различии признаков Quercus от других родов. Во время среднего эоцена также заслуживают внимания репродуктивные структуры, обнаруженные у Quercus oligocenensis в Теннесси. В позднем эоцене секции Quercus, Lobatae и Protobalanus, а также некоторые дубы Cyclobalanopsis присутствовали в Северной Америке, тогда, когда секции Lobatae и Cyclobalanopsis встречались в Европе. С другой стороны, секция Protobalanus появилась в эоцене в Северной Америке и, по-видимому, всегда была ограничена этим континентом (Barron et al., 2017).
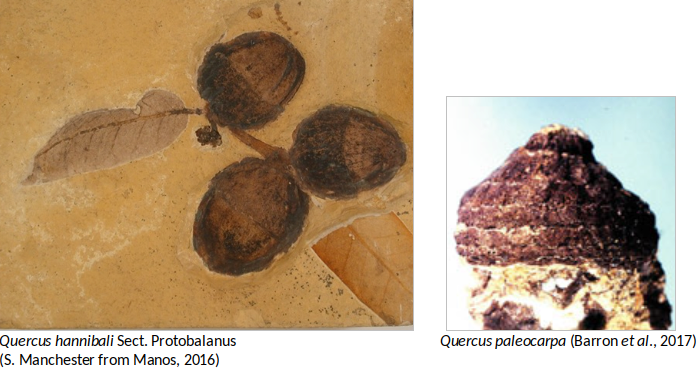
Листья , деревисина, пыльца, плоды и цветы, принадлежащие к видам Quercus, распространены от олигоцена до четвертичной флоры Северного полушария. Из-за такого обилия листьев в окаменелостях дубов идентификация часто основана на одном или нескольких экземплярах, затрудняющих различение близкородственных видов Quercus. По этим причинам текущее количество ископаемых видов дубов, скорее всего, преувеличено и нуждается в серьезном пересмотре. Примечательно, что единственное упоминание о группе Cerris в Америке происходит с этого времени, представленной остатками листьев Quercus prevariabilis (Barron et al., 2017). Желуди от миоценовой флоры распространены на западе Северной Америки, как Quercus hannibali (раздел Protobalanus) из Невады. Хотя эти желуди очень распространены, обычно они не очень хорошо сохранились, что затрудняет их идентификацию (Barron et al., 2017). Запись неогена (миоцена и плиоцена) представлена в основном окаменелостями пыльцы в восточной части Северной Америки и показывает появление и расширение ареала секций Quercus, Lobatae и Protobalanus.
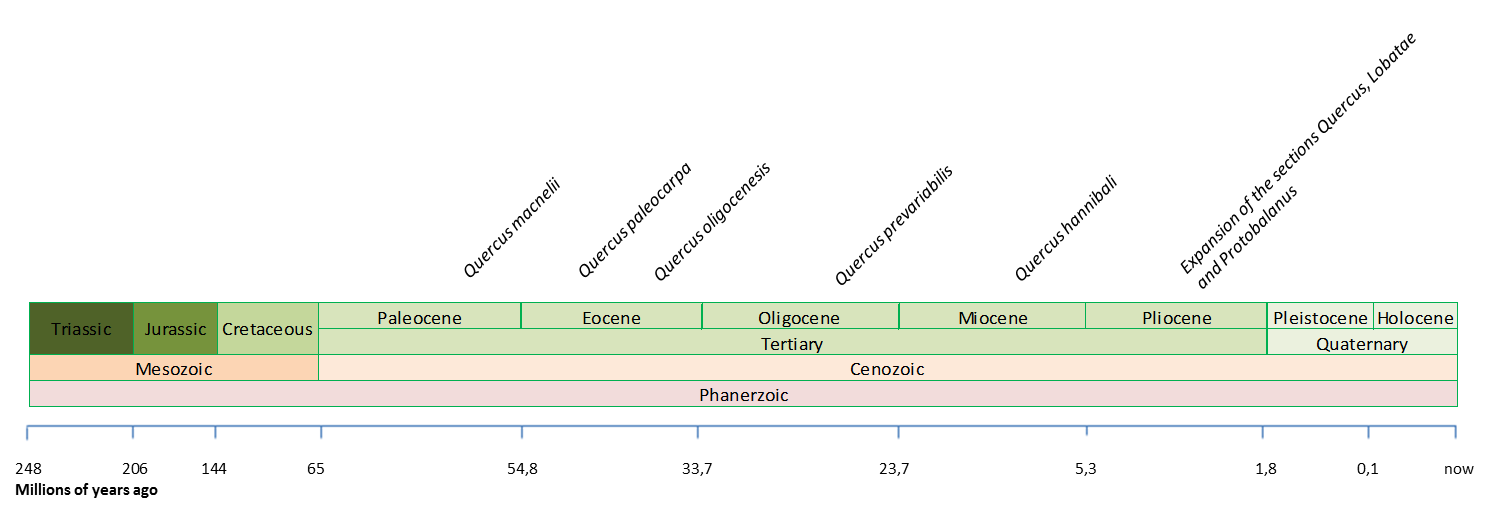
Наиболее важные найденные окаменелости рода Quercus в Америке (Barron et al., 2017; Xu et al., 2014; Binh et al., 2018. Иллюстрация Zamira Betancourt).
ЭВОЛЮЦИОННАЯ БИОЛОГИЯ ПОЗДНЕГО ПЛЕЙСТОЦЕНА
В течение четвертичной периода дубы в северном полушарии подвергались важным миграциям в ответ на климатические изменения. Было около 17 климатических колебаний (чередование ледниково-межледниковых периодов), в течение которых виды ареалы дубов то расширялись, то уменьшались. Ледниковый период длился от 50 до 100 тысяч лет, тогда как межледниковые периоды были значительно короче и длились от 10 до 20 тысяч лет. Климатические колебания были сильными избирательными силами, предпочитающими те виды, которые были достаточно подвижны, чтобы продвигаться в более благоприятные места обитания (Dynesius and Jansson, 2000). Скорее всего, они были ответственны за сокращение числа видов, которые занимают большое континентальное распространение (Q. robur в Европе, Q. alba в Америке или Q. acutissima в Азии). Эти движения оказали сильное влияние на генетическое разнообразие видов, но в Северной Америке и Европе довольно разными путями (Grivet et al., 2006; Kremer et al., 2010). Благодаря объединению палеоботанических и филогеографических исследований пути миграции с момента последнего ледникового максимума были реконструированы в Европе и Северной Америке.
В Европе
Большое исследование, проведенное в Европе, сопоставляющее оставшиеся исторические следы (отложения пыльцы) с генетическими отпечатками пальцев (полиморфизмы ДНК хлоропластов (cpDNA)), продемонстрировало, как существующее распределение генетического разнообразия было сформировано динамикой постледниковой колонизации (Kremer, 2002). В конце последних оледенений европейские дубы были ограничены тремя основными убежищами (Южный Пиренейский полуостров, Центральная Италия и Южные Балканы).
Поскольку ледниковые периоды длились более 100 000 лет, виды, по всей видимости, генетически дифференцированы между этими рефугиальными зонами (убежищами во время неблагоприятных условий), о чем свидетельствуют совершенно разные линии гаплотипов (вариант cpDNA), занимающие эти районы. Менее чем за 7000 лет (от 13 000 до 6 000 лет до н.э.) дубы реколонизировали большинство своих современных ареалов, начиная с районов убежища (Petit et al., 2002a и b; Brewer et al., 2002). В горных районах (Пиренеи, Юго-Восточные Альпы и Карпаты) от 13 000-10 000 до н.э. количество дубов резко возросло.
Снижение температуры в течение от 11 000 до 10 000 лет остановило это расселение и привело к сокращению численности. После 10 000 лет до н.э дубы распространились по всей Европе и достигли своего распространения примерно в 6 000 лет до н.э. Расширение было более быстрым на западе и более медленным в центре и на востоке из-за Альп и Карпатских гор. В среднем миграция была чрезвычайно быстрой (от 300 до 500 метров в год) (Brewer et al., 2002). Редкие случаи рассеивания на больших расстояниях внесли значительный вклад в быстрое распространение вида (Le Corre et al, 1997; Davies et al, 2004). Эта динамика колонизации привела к сильной филогеографической структуре европейских белых дубов, где материнские линии современного дубового леса демонстрируют сильное распределение Восток-Запад, что свидетельствует о путях постледниковой миграции (см. Карты).лет. Динамика колонизации также имела противоположные последствия для генетического разнообразия вида. Несмотря на сильный эффект основателя, сопровождавший реколонизацию, дубы смогли сохранить свое генетическое разнообразие. Хотя наибольшее разнообразие ограничено южными районами Европы, уровень разнообразия по-прежнему высок и в центральной части Европы, где объединены различные миграционные волны, происходящие из зон убежища (Petit et al., 2002a).
Однако сегодняшнее распределение адаптивного разнообразия не связано с первоначальным разнообразием. Географическая изменчивость адаптивных черт обусловлена более поздним локальным давлением отбора. Межвидовая гибридизация была ключевым механизмом миграции, так как она способствовала распространению поздних сукцессионных видов (Q. petraea) в пионерские виды (Q. robur). Систематическое совместное использование одного и того же гаплотипа кДДНК различными видами белого дуба, занимающими одинаковые насаждения, указывает на то, что гибридизация была обширной во время послеледниковой реколонизации (Petit et al., 1997).
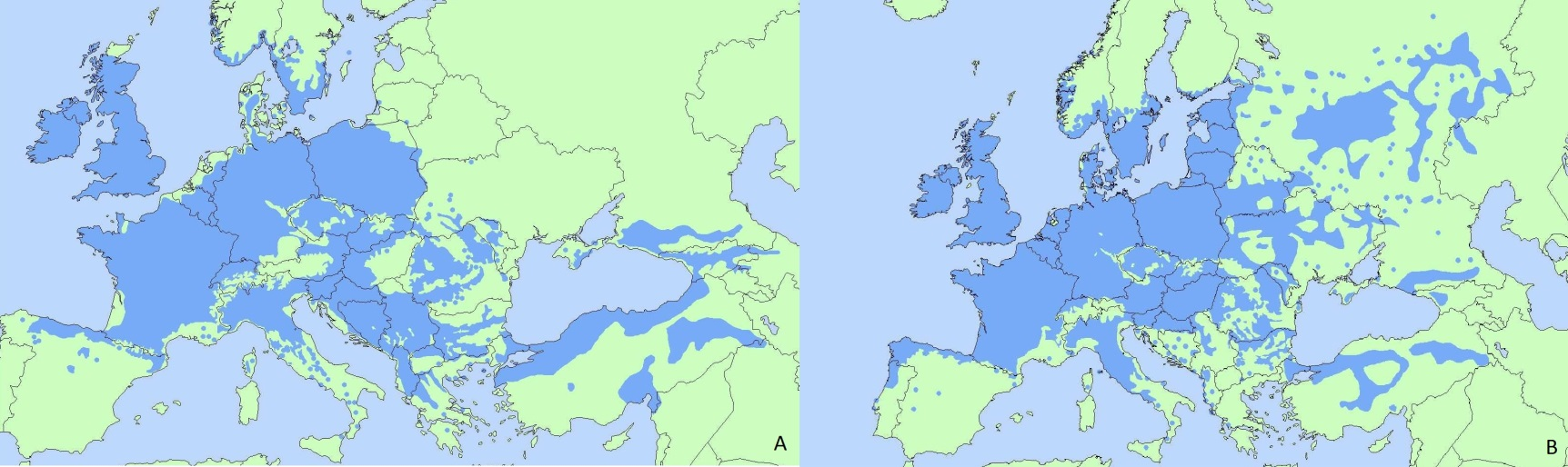
Текущее распространение Quercus petraea (A) и Quercus robur (B).
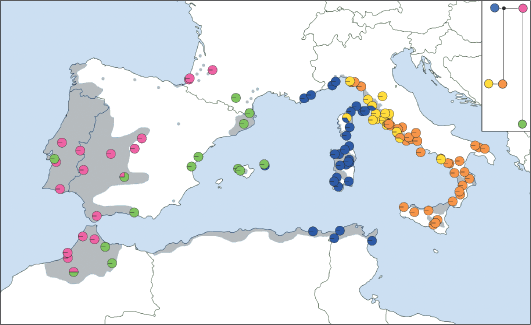
Распределение гаплотипов кПДНК в популяциях Quercus suber и филогенетическая реконструкция отношений между гаплотипами. Серая зона соответствует современному распространению Q. suber. (Magri et al., 2007).
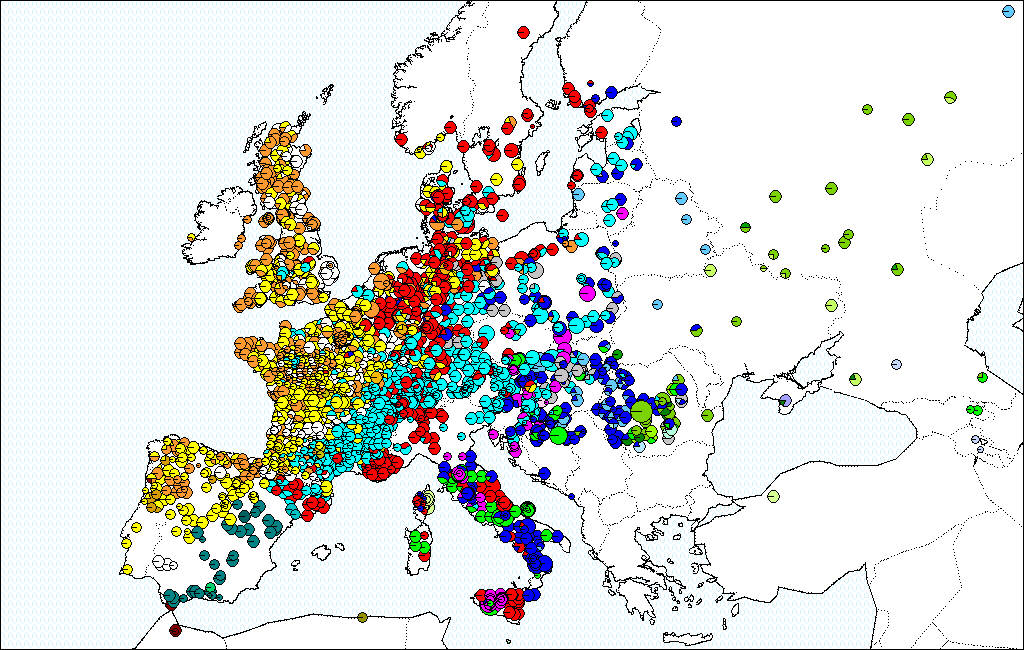
Распределение гаплотипов (отдельных генетических вариаций) у европейских белых дубов, представляющих 42 гаплотипа в более чем 3000 популяциях (Petit et al., 2002).
В северной Америке
Динамика постледниковой колонизации в Северной Америке сильно отличалась от европейской. С восточной стороны виды не были ограничены генетически отдельными рефугальными зонами. Кроме того, дубы сохранились в виде популяций с низкой плотностью, близких к Лаврентидному ледниковому щиту, уменьшая вероятность для рассеивания на большие расстояния. Следовательно, послеледниковая реколонизация была более диффузной, чем в Европе, и была ограниченной (Schlarbaum et al., 1982). В результате дубы восточной части Северной Америки демонстрируют меньшую дифференциацию кДДНК среди своих популяций (Magni et al., 2005).
С западной стороны дубы также были более устойчивыми в ответ на климатические колебания. Калифорнийские дубы не вымерли полностью в течение последнего ледникового периода. Популяции уменьшились в размерах и расширились в периоды потепления. В это же время локальное перемещение отдельных видов приводит к неравномерному распределению разнообразия кПДНК и поддерживает более высокие уровни разнообразия по сравнению с европейскими дубами (Grivet et al., 2006; Dodd and Kashani, 2003) , Несколько похожая картина была описана в случае со средиземноморскими дубами в Испании, что свидетельствует о сохранении многих популяций, которые сохранились до сих пор и сохраняют большой уровень разнообразия (De Heredia et al, 2007).
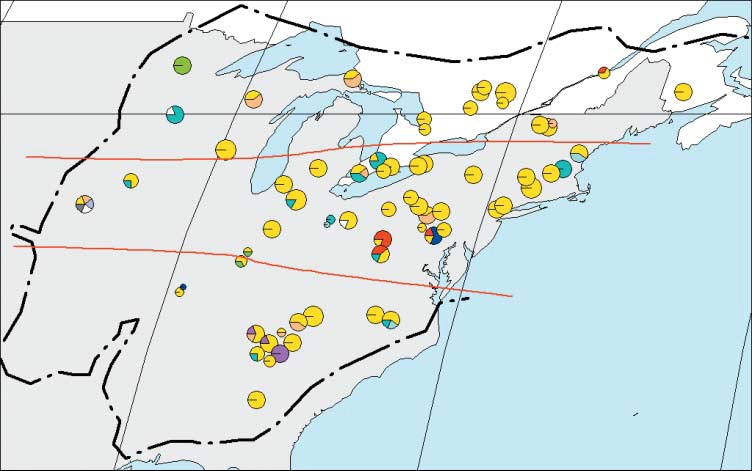
Географическое распределение и частота гаплотипов в каждом участке во всем естественном ареале Quercus rubra (пунктирная линия). Красные линии показывают три широтных подразделения. B. Цвета гаплотипов и филогенетические связи между гаплотипами Quercus.rubra, обнаруженные с помощью ПЦР-RFLP.
